
Pemuliaan tanaman dengan teknik seleksi dan
hibridisasi memiliki banyak keterbatasan, terutama karena ketidakmampuan teknik
ini untuk menembus barrier reproduksi,
sehingga sifat yang diperoleh berasal dari spesies itu sendiri. Untuk itu,
muncullah teknologi transgenik dengan memanfaatkan DNA rekombinan yang kemudian
ditransformasikan ke dalam sel tanaman yang selanjutnya dikultur menjadi
tanaman transgenik yang utuh dan baru. Untuk menghasilkan tanaman transgenik,
ada beberapa langkah utama yang dilakukan, yaitu:
1.
Isolasi gen
2.
Menyiapkan jaringan
(kultur jaringan)
3.
Transformasi
4.
Regenerasi
Pada artikel ini, pembahasan akan lebih ditekankan
pada tahap transformasi vektor yang membawa gen ke dalam sel tanaman.
Metode Transformasi Genetik pada Tanaman
Ada beberapa metode yang dapat digunakan untuk
transformsi gen ke dalam tanaman, dan yang umum digunakan yaitu partikel bombardment dan melalui pertantara
Agrobacterium tumefaciens.
1. Partikel
Bombardment
 |
| Helios Gen Gun dan alat Gen Gun Standar |
Partikel bombardment merupakan salah satu teknik yang
digunakan untuk memasukkan DNA asing ke dalam kultur sel dengan cara
penembakan. Melalui teknik ini, gen asing ditransfer secara langsung ke dalam
sel atau jaringan, dengan daerah kisaran yang luas. Partikel Bombardmen juga
dikenal sebagai gen gun (senapan gen) atau biolistik (Anonim, 2009). Teknik partikel
bombardent sekarang sudah luas digunakan, tidak hanya untuk produksi tanaman
transgenik tetapi juga digunakan pada sel bakteri dan hewan bahkan ke dalam
organ hewan yang masih hidup. Para saintis telah memodifikasi alat ini sehingga
dapat mentransfer DNA ke dalam mitokondria yeast, kloroplas dan mikroalga
hijau (Clark, 2012: 406).
Gen terkandung dalam plasmid ataupun dalam kaset gen (fragmen
linear gen hasil PCR). Pada plasmid terdapat marker khusus, promoter, sekuen
gen dan terminator sedangkan kaset gen terdiri atas promoter, primer, probe,
sekuen gen, terminator. Transformasi menggunakan partikel bombardment dapat
dilakukan dengan lebih dari satu plamid yang membawa gen berbeda untuk
berintegrasi dalam genom tumbuhan yang sama, contohnya gen yang mengkode PHA
untuk jalur metabolit (Romano, dkk,. 2003).
 |
| Skema Kaset gen |
Teknik
kerja partikel bombardment
Pada teknik bombardment, DNA dibawa oleh partikel
logam emas atau tungsten yang berukuran mikroskopik. Partikel yang digunakan
untuk bombardment biasanya berupa emas karena bersifat inert, padat, dan tidak
beracun dalam sel. Partikel emas lebih aman untuk digunakan karena tungsten
dapat bersifat toksik pada beberapa tanaman. Partikel yang membawa DNA ditembakkan oleh senapan (gun) ke dalam
jaringan tanaman dan menembus dinding sel untuk masuk ke dalam nukleus,
mitokondria atau kloroplas dan menyatu dengan kromosom DNA inang. DNA
vektor/plasmid membawa sekuen spesifik yang digunakan untuk mengenali lokasi
yang tepat atau yang diinginkan untuk berintegrasi dengan genome (Hayes dan
Frieda, 2010).
Clark (2012) menyebutkan bahwa gen gun dapat
beroperasi melalui 2 cara yaitu dengan pemberian tekanan udara maupun tegangan
voltase yang tinggi.
 |
| Tipe Gen Gun. A. tekanan udara; B. tegangan voltase tinggi |
DNA melepaskan diri dari partikel di dalam sel.
Beberapa DNA masuk ke dalam organel target dan berhasil berintegrasi dengan
kromosom DNA inang. Kemudian sel/jaringan transgen ditumbuhkan dalam medium
kultur kusus sesuai dengan DNA yang membawa penanda kusus, misalnya gen
resisten herbisida atau insektisida, gen yang dapat mendegradasi mercuri. Hasil
kultur dianalisis untuk mengetahui hasil ekspresi DNA asing. Teknik analisis
untuk mengetahui ekspresi transgen dilakukan melalui analisis sourthen blotting
atau PCR (Romano, dkk., 2003).
Faktor-faktor
keberhasilan
Ada beberapa variabel yang harus dikontrol agar
tingkat keberhasilan transformasi efektif, meliputi
- Temperatur, jumlah sel dan kemampuan regenerasi
sel atau totipotensi yang digunakan. Eksplant yang digunakan lebih baik
eksplant yang masih memiliki kemampuan mersitematis, misalnya jaringan
embrional dan epikotil (Indurker,
2006). .
- Jumlah DNA yang menyelubungi partikel logam yang
ditransfer ke dalam sel/jaringan (Eisenbraun, 1993)
- Tipe senapan yang digunakan, jenis microcarrier,
pemberian tekanan helium (Indurker,
2006).
Keuntungan
- Gen gun dapat digunakan pada jangkauan yang lebih
luas, misalnya pada tumbuhan dapat digunakan organ daun. Bakteri atau
virus tidak dapat mentransfer gen ke dalam kloroplas sehingga metode gen
gun dapat digunakan untuk memasukkan DNA asing ke dalam kloroplas.
- Transformasi genetik lebih sederhana, cepat, dan
memberikan frekuensi hasil transforman yang lebih tinggi dibandingkan
menggunakan Agrobacterium.
Transformasi menggunakan Agrobacterium pada buncis memiliki frekuensi
hasil transformasi rata-rata 0.5-3%, sedangkan menggunakan partikel
bombardment yaitu 18% (Indurker, 2006).
Keterbatasan
Keterbatasan transformasi menggunakan partikel
bombardment dibandingkan Agrobacterium yaitu
- Harga alat dan perlengkapannya cukup mahal
- Penembusan partikel ke dalam jaringan cukup
dangkal
- Daerah penembusan DNA plasmid ke dalam jaringan
target cukup luas sehingga DNA yang ditransfer ke dalam sel acak. Beberapa
sel yang tidak mengekspresikan transgen, akan mati jika ditumbuhkan dalam
medium kultur kusus (Clark, 2012: 405).
- Seringkali terjadi penggabungan salinan ganda
transgen pada sisi tunggal penyisipan, penyusunan ulang gen yang menyisip
dan penggabungan transgen pada sisi penyisipan ganda. Salinan ganda dapat
menyebabkan hilangnya transgen pada keturunan berikutnya (Yao dkk., 2006).
- Salah satu permasalahan dalam semua metodologi
transformasi yang digunakan dan pada penggunaan partikel bombardment
sendiri adalah tidak ada jaminan terekspresinya transgen oleh tanaman.
Epigenetik dan efek posisi, gen silencing, fenomena supresi dan co-supresi
seringkali menginaktifkan transgen (Romano, dkk., 2003).
2. Agrobacterium
tumefaciens
Agrobacterium
adalah genus dari bakteri gram negatif yang ditemukan oleh H.J.Conn yang
digunakan untuk transfer gen secara horizontal yang menyebabkan tumor. Agrobacterium tumefaciens adalah bakteri patogen
pada tanaman yang banyak digunakan untuk memasukkan gen asing ke dalam sel
tanaman untuk menghasilkan suatu tanaman transgenik.
Secara alami, A. tumefaciens dapat menginfeksi tanaman dikotil
melalui bagian tanaman yang terluka sehingga menyebabkan crown gall tumor.
Bakteri yang tergolong ke dalam gram negatif
ini memiliki sebuah plasmid
besar (lebih dari 200 kb) yang disebut plasmid-Ti
yang berisi gen penyandi faktor virulensi
penyebab infeksi bakteri
ini pada tanaman. Untuk memulai pembentukan tumor, A. tumefaciens harus
menempel terlebih dahulu pada permukaan sel inang dengan memanfaatkan polisakarida
asam yang akan digunakan untuk melakukan
kolonisasi pada sel tanaman. Selain
tanaman dikotil, tanaman monokotilseperti
jagung,
gandum,
dan tebu
telah digunakan untuk memasukkan sel asing ke dalam genom tanaman.
 |
| Tumor yang disebabkan oleh Agrobacterium |
Plasmid Ti adalah vektor
alami yang digunakan untuk mentransfer DNA
ke dalam sel
tanaman.
Pada sebagian besar plasmid Ti, terdapat empat
kompleks gen, yaitu T-DNA (bagian yang ditransfer dan menyatu dengan genom
tanaman, gen virulen (vir) yang
terdiri dari 50 kilo-basa untuk mengatur proses transfer T-DNA ke dalam DNA
tanaman, gen tra/trb yang mengatur perpindahan plasmid Ti antarbakteri
(conjugative transfer), bagian yang mengatur sistem replikasi
plasmid
(ORI), dan bagian gen
yang menyandikan katabolisme opine. Molekul opin ini akan dihasilkan
oleh jaringan
tanaman
yang terinfeksi bakteri
pembawa plasmid Ti dapat berupa octopine,
nopaline,
succinamopine
and leucinopine.
Plasmid Ti ini memiliki 196 gen yang dikode oleh 195 protein, memiliki panjang
206,479 nukleotida, kandungan GC 56% dan 81% material yang dikode oleh gen.
 |
| Peta Ti plasmid Agrobacterium tumefaciens |
Daerah
virulensi (virulence region) terdiri gen virABCDEFG yang mengkode suatu enzim yang bertanggung
jawab untuk mentransfer T-DNA ke dalam sel tumbuhan, yaitu
· virA mengkode reseptor (transmembrane
dimeric sensor protein) yang beraksi
ketika adanya senyawa phenolic berupa acetosyringone, syringealdehyde
atau acetovanillone yang dikeluarkan dari
kerusakan jaringan tumbuhan.
·
virB mengkode protein yang menghasilkan struktur seperti pilus
·
virC berikatan dengan enhancer pada T-region
·
virD1 dan virD2 mengenali T-DNA border dan menghasilkan endonuklease
yang memotong (nicking) ujung kiri dan ujung kanan dari T-DNA yang dimulai dari
ujung kanan
·
virG adalah faktor transkripsi (trancriptional factor) yang
mengaktifkan ekspresi gen Vir setelah berikatan dengan sekuens yang cocok.

Pada
kromosom Agrobacterium setiap elemen
gen menunjukkan peranan yang berbeda untuk perlekatan A. Tumefaciens ke sel tumbuhan. Lokus chvA dan chvB
terlibat dalam sintesis dan ekskresi β 1,2 glucan (Cangelosi et al.,
1989), chvE
dibutuhkan dalam pengenalan gula dari induksi gen vir dan untuk kemotaksis
bakteri tersebut (Ankenbauer et al.,
1990, Cangelosi et al.,
1991), lokus cel bertanggung jawab untuk sintesis fibril selulosa (Matthysse 1983),
lokus pscA (exoC) berperan dalam siklus glucan dan asam
sukkinoglikan (Cangelosi et at.,
1991), dan lokus att yang terlibat dalam pembentukan protein permukaan sel bakteri
(Matthysse, 1987).
Di
alam, Agrobacterium tertarik pada
tumbuhan yang memiliki luka kecil yang mengeluarkan senyawa phenolik seperti
acetosyringone dan gula. Senyawa ini menginduksi bakteri untuk berpindah dan
melekat pada tumbuhan melalui berbagai macam receptor permukaan sel. Induser
yang sama mengaktifkan ekpresi gen vir yang terdapat pada Ti plasmid yang
bertanggung jawab untuk transfer ss- DNA menuju sel tumbuhan. Ini di bawah kendali dua komponen sistem
regulasi yaitu virA protein yang mengenali acetosyringone yang dikode oleh gen
virA di dalam Ti-plasmid dan chvE protein yang mengenali gula yang dikode di
dalam kromosom bakteri . Senyawa-senyawa tersebut mengeluarkan signal yang
dikenali oleh reseptor dimeric transmembran kompleks virA chvE. Pada permukaan
sel, sensor melakukan aktivasi Vir A dengan cara autophosphorilasi ketika
mendeteksi senyawa phenolic tumbuhan. Selanjutnya Vir A akan mengirimkan
phosfat untuk pengikatan DNA oleh protein Vir G sebagai faktor transkripsi yang
mengaktifkan proses transkripsi gen vir pada
plasmid Ti yang akan mengekpresikan virC,
virD, virE, virB, virF dan virH. Dua gen yang dihasilkan VirD1 dan VirD2 yang mengenali 25 pb pada
kedua ujung T-DNA yang kemudian
memotongnya (nicking) membentuk kompleks
untai tunggal T yang belum matang (immmature) yang disebut kompleks
ssT-DNA-VirD2 . secara in vitro membuktikan
bahwa kehadiran dari virD1 sangat dibutuhkan untuk memotong ssT-DNAoleh virD2.
VirD2 pada saat itu melekat pada ujung 5’ akhir dari T-DNA dan memotongnya
secara endonukleolitik sehingga akan membentuk gap (celah), dan helikase
bakteri melepaskan T-DNA dari plasmid. Celah (gap) untai tunggal pada plasmid tersebut akan
segera diperbaiki. Kemudian T-DNA akan ditempatkan pada suatu cekungan yang
diselubungi dengan protein VirE2 yang
disebut dengan hollow cylindrical
filament dengan struktur yang bergulung. Ini adalah bentuk matang (mature)
dari T-DNA yang siap masuk ke dalam sel tumbuhan.
Sebenarnya ada dua model teori
pengiriman kompleks ssT-DNA-VirD2 yang
telah dikemukakan. Tetapi, yang banyak diterima adalah model penyelubungan
untai tunggal oleh protein VirE2 (single strand binding protein virE2) dan
mesin transfer (virB) kemungkinan tidak berinteraksi secara langsung dengan
T-DNA. Pada alternatif model yang kedua,
kompleks ssT-DNA-VirD2 nampak
telanjang karena tidak diselubungi oleh
virE2 sehingga terjadi interaksi langsung antara mesin transfer (virB) dengan
kompleks ssT-DNA-VirD2, sedangkan virE2 ditansfer secara independent oleh mesin
transfer ke dalam sel tumbuhan. Telah diketahui bahwa virE1 sangat dieprlukan
untuk ekspor virE2 kedalam sel tumbuhan. Strain bakteri yang telah dimutasi
virE1 nya tidak dapat mengekspor virE2 sehingga terakumulasi di dalam sel
bakteri tersebut. T-DNA ditansfer ke tumbuhan sama halnya dengan konjugasi
bakteri. Pertama-tama Agrobacterium
membentuk suatu pilus yang merupakan ekpresi dari gen virB. Pilus ini
menyerupai batang yang menghubungan dengan sel tumbuhan dan membuka saluran
yang siap ditansferkan secara aktif T-DNA ke dalam sitoplasma tumbuhan. Pilus
dan kompleks transport terdiri dari protein yang dihasilkan oleh gen vir.
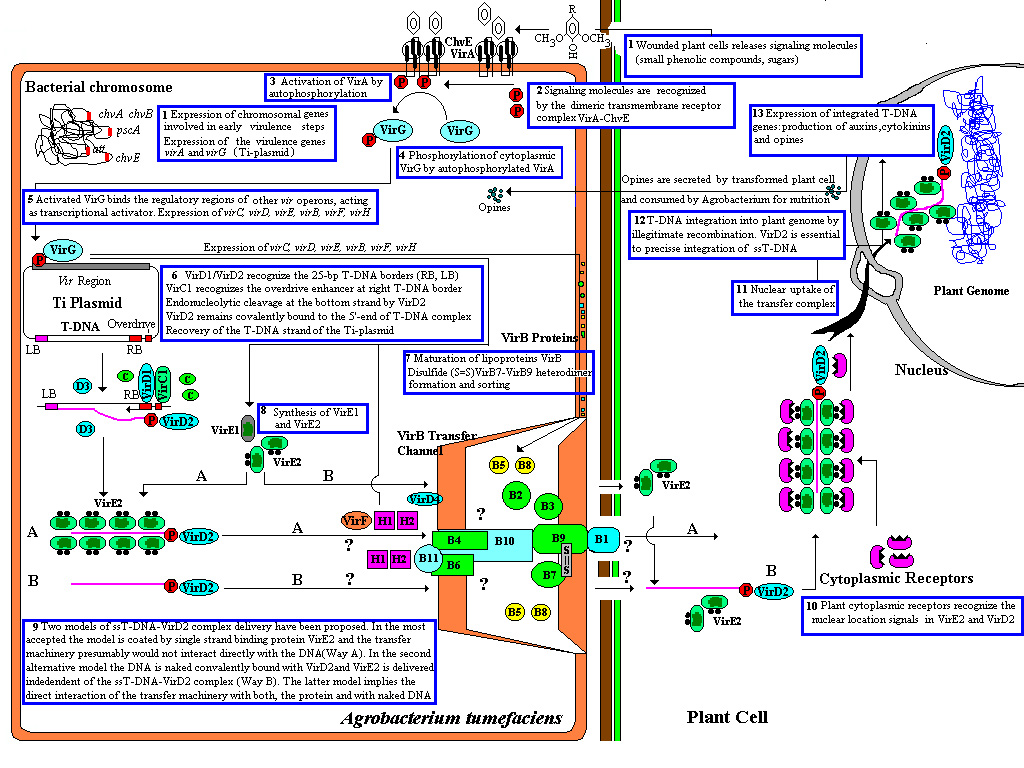
Proses transfer T-DNA Agrobacterium
tumefaciens
Gen
pada T-DNA akan diekspresikan sama halnya pada eukaryot yang memiliki
promoters, enhancer dan bagian poly (A). Oleh sebab itu, ekspresi dalam nukleus tumbuhan lebih baik
dibandingkan pada Agrobacterium.
Protein ini akan menyandi sintesis dua hormon pertumbuhan yang auksin dan
sitokinin. Auksin membuat sel tumbuhan menjadi lebih besar dan sitokinin
berperan dalam pembelahan sel. Sel tumbuhan yang diinfeksikan ini akan memulai
tumbuh cepat dan tanpa kontrol sehingga menghasilkan tumor.
T-DNA
juga membawa gen untuk mensintesis opine yang mana merupakan variasi yang
berbeda dari asam amino dan derivat gula fosfat. Opine dihasilkan oleh sel
tumbuhan yang dikandung T-DNA tetapi digunakan oleh bakteri sebagai sumber
carbon, nitorgen dan energi. Ini adalah
cara bagaimana bakteri menggunakan tumbuhan untuk menghasilkan sumber makanan
bagi bacteri. Plasmid Ti selalu berada dalam Agrobacterium, membawa gen yang menyediakan bakteri untuk
mendapatkan opin.
Dalam
prakteknya, Agrobacterium digunakan
untuk mentransfer gen dari suatu kepentingan kedalam tumbuhan menggunakan
kultur jaringan. Tiap pemisahan sel tumbuhan disebut protoplas atau sebuah bagian
dari kalus yang di kultur dengan Agrobacterium
mengandung sebuah plasmid Ti yang dimodifikasi T-DNA nya. Setelah kokultur, sel tumbuhan dipanen dan di inkubasi dengan
herbisida dan antibiotik yang digunakan sebagai marker selektif. Ini akan
membunuh semua sel yang tidak ditransformasikan T-DNA atau gagal untuk
mengekspresikan gen pada T-DNA. Sel yang telah ditransformasikan dapat di
induksi untuk menghasilkan tunas dan jaringan akar dengan mengubah kondisi
hormon pada medium mudah diuraikan. Tumbuhan transgenik yang masih kecil dapat
dilindungi untuk level ekspresi transgen berikutnya.
Daerah
T-DNA dari Ti plasmid dapat direkayasa genetika dengan menambah gen resisten
antibiotik (antiobiotic resistance gene
(kanR)) dan
DNA asing yang diinginkan. Integrasi DNA asing kedalam sel tumbuhan mengganggu
pembentukan tumor dan hanya sel tumbuhan dengan gen kanR yang dapat tumbuh pada kultur yang mengandung
antibiotik. Tumbuhan sangat mudah beregenerasi dari kultur sel (kalus) dan
tumbuhan transgenik yang telah dewasa mengekspresikan gen asing.
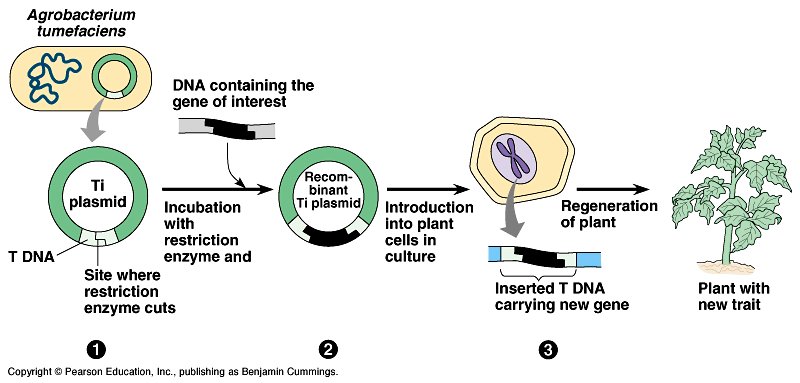
Produksi tumbuhan transgenik dengan menggunakan integrasi Ti plasmid.
Baru-baru
ini, sebuah metode yang disebut dengan in planta Agrobacterium transformation telah
dikembangkan dan merevolusi dunia transformasi tumbuhan. Transformasi in planta juga diketahui sebagai metode
floral dip. Metode ini telah dikembangkan menggunakan tumbuhan model Arabidopsis tetapi sedang diperluas
untuk tumbuhan lain, seperti gandum dan jagung (Gambar 16): A. Pertama, Arabidopsis ditumbuhkan sampai tunas bunga mulai terbentuk. Tunas
kemudian dipindahkan dan dibiarkan beregenerasi untuk beberapa hari. B. Ketika mulai
beregenerasi, tumbuhan dicelupkan ke dalam suspensi Agrobacterium yang berupa surfaktan. Surfaktan Agrobacterium dibiarkan
untuk melekat pada tumbuhan dan mentransfer T-DNA nya. Karena tunas bunga sudah
mulai terbentuk, T-DNA akan menjadi bagian dari jaringan ovarium sampai
akhirnya tumbuhan menyelesaikan pertumbuhan dan pembentukan bibitnya. C. Tanaman dipelihara selama beberapa minggu hingga
dewasa dan kemudian bibit anakan dipanen. D.
Bibit
tersebut dipanen dan ditumbuhkan pada medium selektif untuk mendapatkan gen
yang terintegrasi dan ekpresi T-DNA. Meskipun, metode ini memberikan persentase
rendah terbentuknya transforman.
 |
| Metode in planta Agrobacterium transformation |
Agrobacterium
merupakan sistem transformasi gen yang menguntungkan karena efisiensinya tinggi
dan integrasinya stabil. Agrobacterium tumefaciens dinyatakan
dapat membawa setiap gen yang diinginkan di dalam T-kompleks dan memasukkannya
ke dalam DNA target pada tanaman dengan tingkat keberhasilan yang tinggi. Hal tersebut dikarenakan untai T-DNA Agrobacterium tumefaciens tidak seperti
komponen genetik mobile pada transposon dan retrovirus yang menyandikan fungsi
bagi pergerakan dan integrasi DNA.
Transformasi
dengan Agrobacterium juga memiliki beberapa keuntungan lain, diantaranya
bersifat dapat diulang (reproducible), relatif lebih murah, memberikan
pola integrasi yang tegas, jumlah salinan dalam genom sedikit (1-3 salinan)
sehingga memudahkan untuk membedakan sifat ekspresi tanaman transgenik itu
sendiri.. Pada awalnya teknik
transformasi dengan Agrobacterium hanya berhasil pada tanaman dikotil
ketika tanaman ini menghasilkan senyawa induser untuk menginduksi gen vir ketika
tanaman luka dan mengeluarkan getah. Tanaman tembakau dan solanaceae adalah
contoh pertama tanaman dikotil yang berhasil ditransformasi.
Selain
menyisipkan gen target untuk perubahan sifat tanaman tertentu yang dikehandaki,
transformasi genetik dengan Agrobacterium pada tanaman juga bermanfaat
untuk membuat populasi tanaman mutan. Dengan menggunakan Agrobacterium memungkinkan
diperoleh mutan dalam jumlah banyak dalam suatu periode yang relatif singkat.
Pembuatan mutan dilakukan dengan menggunakan elemen loncat (transposon)
misalnya transposon Ac/Ds. Transposon Ds akan berpindah posisi
dalam genom pada tempat berbeda dan tersisip pada gen-gen fungsional. Sedangkan elemen Ac menyandikan suatu
enzim yang mengaktifkan elemen Ds untuk bertransposisi. Adanya
penyisipan Ds ini memungkinan fenotipe tanaman menjadi beragam.
Keragaman mutan ini dapat dijadikan sebagai sumber plasma nutfah baru untuk
selanjutnya dapat dilakukan isolasi gennya (Mulyaningsih, 2009).
Proses
transformasi gen via Agrobacterium
juga dapat dilakukan dengan sistem vektor biner. Sistem vektor biner yang
diterapkan dalam proses transformasi gen via Agrobacterium dapat meningkatkan efisiensi transformasi. Sistem binary vector merupakan
penggunaan dua plasmid Ti secara bersama dan saling berhubungan untuk proses
tranfer gen. Kedua plasmid Ti tersebut terdiri dari satu plasmid pembawa range
replikon yg luas (ORI dari E.coli dan
Agrobacterium tumefaciens, T-DNA yang
mengandung multiple cloning site, gen resistan antibiotik) sedangkan plasmid
pasangannya mengandung gen virulence (vir-region) tanpa T-DNA.
Umumnya Agrobacterium
tumefaciens sebagai media transformasi gen relatif efisien diterapkan pada
spesies tumbuhan. Dilain pihak, ada beberapa spesies tanaman yang tingkat
keberhasilan transformasinya rendah, sebagian besar adalah jenis tanaman monokotil. Namun, menurut Mulyaningsih (2009)
pengembangan penelitian terkini dapat mengatasi kelemahan tersebut dengan melakukan beberapa
penyesuaian kondisi seperti penambahan senyawa induser dan pH saat
ko-kultivasi. Hiei dkk (1994) dalam Mulyaningsih (2009) dalam telah berhasil
membuktikan bahwa tanaman padi jenis japonica berhasil ditransformasi
menggunakan Agrobacterium dengan material tanaman berupa sel kalus
embriogenik. Dalam penelitiannya Hiei dkk menambahkan senyawa asetosiringone
pada media dan menggunakan media dengan pH 5,2 saat ko-kultivasi. Hingga saat
ini studi transformasi genetik dengan Agrobacterium terhadap tanaman
pangan seperti padi terutama jenis indica (yang banyak dibudidayakan dan
dikonsumsi) terus dilakukan. Beberapa
jenis tanaman pangan dan non pangan hasil transformasi dengan Agrobacterium di
Amerika yang dilaporkan ialah kedelai, kapas, jagung, bit, , gandum, canola, creeping
bentgrass (untuk pakan).











0 komentar:
Posting Komentar